Welcome to the Pet Food Knowledge Centre


What We Feed Our Pets Can Affect Their Digestive Health
A healthy digestive system is important for providing a physical and immunological barrier to potential pathogens in the environment and extracting and absorbing nutrients from food to meet the animal’s nutritional requirements. In recent years, it has become increasingly apparent that a healthy microbiome plays a vital role in digestive health and contributes to maintaining overall health and well-being. The term ‘gut microbiome’ refers specifically to the trillions of microorganisms living in the intestinal tract. While some microorganisms are harmful to a pet’s health, many are incredibly beneficial and necessary for a healthy body. Microbes have the capability to unlock and synthesize nutrients that have direct benefits to the pet. Many factors can influence the population of the microbiome, such as age, diet, environment, and antibiotics. Still, diets are often supplemented with ingredients to help promote the growth of healthy gut bacteria to support the best intestinal health. Prebiotics, probiotics, and postbiotics are just a few ingredients that can help maintain a healthy gut microbiome and digestive health.
Prebiotics, Probiotics, and Postbiotics – What’s the Difference?

What are Prebiotics?
Prebiotics have been defined as non-digestible oligosaccharides that stimulate the growth and activity of a limited number of resident colonic bacteria (Gibson and Roberfroid, 1995), which can have a beneficial impact on factors, including digestive health. Two examples of prebiotics are mannan-oligosaccharides (MOS) and fructo-oligosaccharides (FOS). FOS, oligofructose, and inulin are all oligosaccharides that occur naturally in plants, including sugar beet, onion, garlic, asparagus, banana, artichoke, and chicory, and help to maintain healthy gut bacteria.
Mannan-oligosaccharides (MOS) and beta-glucans are prebiotics isolated from yeast cell walls and are collectively termed mannans. MOS are linked to proteins to form a mannoprotein layer localized to the external surface of the cell. MOS are not digested by digestive enzymes in the small intestine and reach the large intestine structurally unchanged. Lactobacilli and some bifidobacteria metabolize MOS and FOS to form short-chain fatty acids (SCFA) – the preferred fuel source of enterocytes, playing a crucial role in supporting the health of the intestinal tract. MOS are less fermentable by intestinal bacteria than fructooligosaccharides (FOS). However, they provide beneficial effects surrounding digestive health, which will be considered in further depth later in this article.
What are Probiotics?
Probiotics are live microorganisms intended to maintain or increase the numbers of “good” bacteria (normal microflora) in the body. Most probiotics do not like elevated temperature, moisture, pressure, and extremes in pH, making it challenging to incorporate them into pet food. The most common microbial species evaluated for probiotics for use in pet foods are Enterococcus faecium and Lactobacillus acidophilus (both lactic acid bacteria). The bacteria use fermentation to produce lactic and acetic acids, which lower intestinal pH and inhibit the growth of certain potentially harmful bacteria. Probiotics present an appealing approach to the treatment and prevention of many conditions because of their potential to be effective and safe and to result in decreased use of medicines.
What are Postbiotics?
Postbiotics are the bioactive compounds and beneficial metabolites made when friendly gut bacteria (probiotics) digest/metabolize/ferment prebiotic substrates. Commercially, postbiotics are produced through precise fermentation processes using specific microorganisms (e.g., yeast) and substrates. According to the International Scientific Association of Probiotics and Prebiotics (ISAPP), a postbiotic is a ‘preparation of inanimate microorganisms and/or their components that confers a health benefit to the host.’ Postbiotics may contain intact inanimate microbial cells and/or fragments with or without metabolites/end products. Postbiotics aim to mimic the beneficial therapeutic effects of probiotics while avoiding the risk and challenges of administering live microorganisms. Consumer interest in postbiotics saw a surge of 91% from 2018 to 2019 (Kerry, 2020). With increasing awareness and focus, it is likely postbiotic claims will increasingly be seen in the pet food and treats market.
How FOS Affects Digestive Health
FOS promotes the growth of friendly gut bacteria such as bifidobacteria and lactobacilli, although some inconsistent results have been observed in different studies. The feeding of dry food with 1% (w/w) oligofructose significantly influenced the fecal bacterial profile in healthy dogs, with increases seen in the numbers of bifidobacteria but also potentially pathogenic species, streptococci, and clostridia (Beynen et al., 2002). Swanson et al. (2002a) reported the results of two studies, each carried out with 20 dogs. In the first study, FOS supplementation resulted in no significant changes in any of the fecal microbial populations evaluated. In contrast, in the second study, a significant increase in bifidobacteria and a non-significant increase in lactobacilli populations were seen. The reason for these differences is unclear, as the only difference between the studies was that the dogs in the first study were slightly older and slightly heavier than the dogs in the second study.
In another study in dogs, changes in fecal bifidobacteria numbers by dietary FOS supplementation were influenced by the protein content of the diet, with a decrease in bifidobacteria seen in dogs fed a ‘low’ protein diet and an increase in bifidobacteria seen in dogs fed a ‘high’ protein diet (Pinna et al., 2018). Regardless of the protein content of the diet, FOS supplementation increased the apparent total tract digestibility of several minerals (Ca, Mg, Na, Zn, and Fe; Pinna et al., 2018). Similarly, Beynen et al. (2002) reported significantly increased magnesium and calcium absorption in dogs fed an oligofructose-supplemented diet. A possible mechanism of action for the increased mineral absorption is that a decrease in ileal pH (i.e., an increase in acidification) raises the solubility of the minerals, making them more available for absorption by the small intestine.
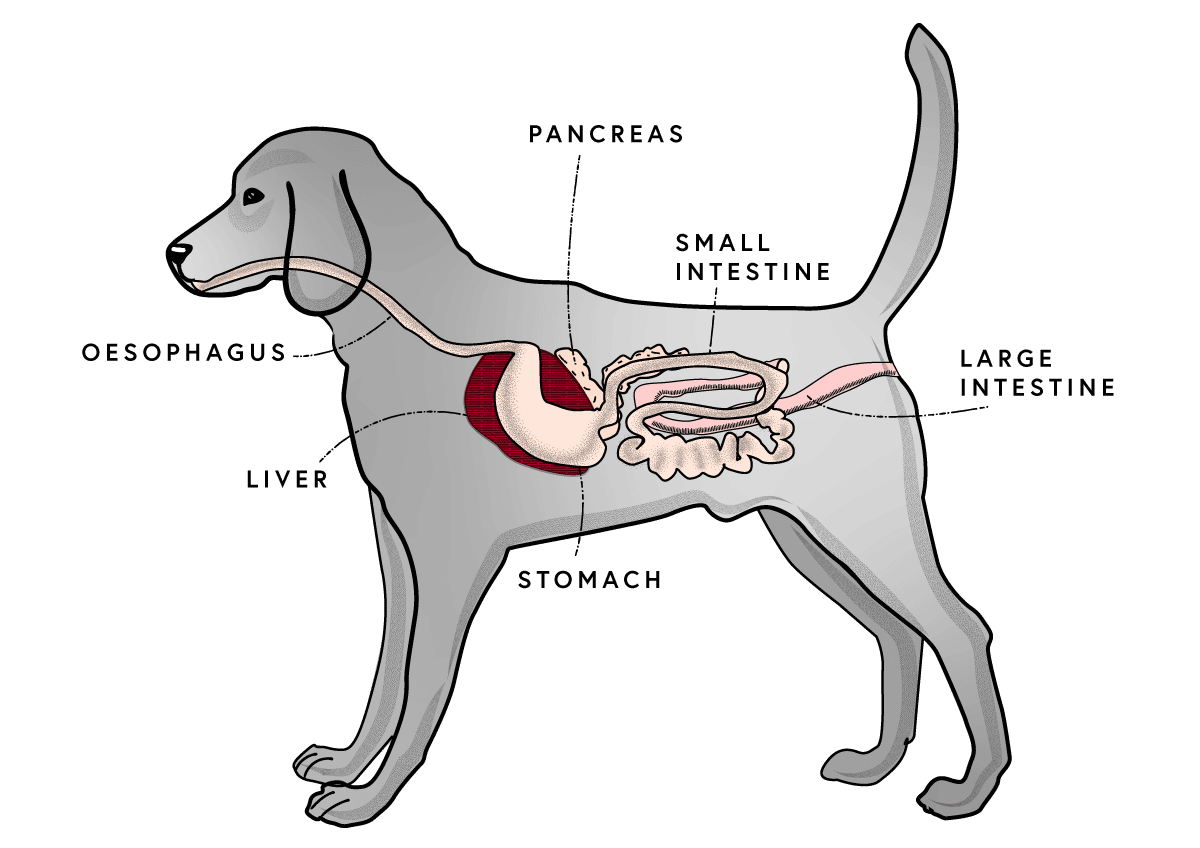
Dietary FOS cannot be digested by the small intestine and reach the large intestine structurally unchanged, where they are metabolized by the intestinal microflora to form short-chain fatty acids. The short-chain fatty acids produced by this process in the gastrointestinal tract stimulate mucosal growth and epithelial cell proliferation within the small intestine (Thompson et al., 1996). Maintaining a healthy colonic mucosa is important to ensure nutrients are absorbed properly and a healthy gut barrier function is maintained. A number of studies have shown that dietary FOS/oligofructose supplementation results in an increase in fecal levels of short-chain fatty acids (acetate, propionate, and butyrate) in dogs (Swanson et al., 2002b; Propst et al., 2003) and increased fecal butyrate in cats (Barry et al., 2010).
A study conducted by Barry and colleagues (2010) suggests that both FOS and pectin were effective fiber sources in promoting intestinal health in cats. Moreover, FOS had increased benefits when compared to pectin because the fructans appeared to produce a more beneficial microbial population than pectin. The study also concluded that the supplementation of fermentable fibers at 4% of a cat diet is successful in modifying stool protein catabolite and microbial concentrations.
How MOS Affects Digestive Health
A study conducted by Grieshop and colleagues (2004) into the gastrointestinal and immunological responses of senior dogs to chicory and mannan-oligosaccharides suggests that MOS and chicory alter fecal microbial populations and certain indices of the immune system. Thirty-four senior dogs were randomly allotted supplements of either 1% chicory, 1% MOS, 1% chicory and 1% MOS, or no supplementation for a 4-week baseline period, followed by a 4-week treatment period. An increase in food intake was noted in dietary supplementation with MOS or MOS and chicory, and this was due to an increase in fermentable fiber and a decrease in the energy content of the diet. Chicory supplementation was seen to increase fat digestibility, and chicory or MOS increased fecal bifidobacteria concentrations, while MOS decreased fecal E. coli concentrations.
A study designed by Kore and colleagues (2012) to assess the effect of dietary supplementation of MOS on nutrient digestibility, hindgut health indices, and plasma metabolic profile found that supplementation of MOS at 1% of diet dry matter positively influenced feed intake, fiber digestibility, and markers of hindgut health. The study used five adult dogs in a complete crossover design. The dogs were fed accordingly on a homemade diet alone or supplemented with MOS (at 1% level). A digestion trial conducted at the end of each period revealed that the intake of dry feed matter and other nutrients increased when supplemented with MOS. The digestibility of fiber was improved in the MOS-supplemented group, while that of other nutrients was not affected. The higher fecal concentration of total SCFAs due to MOS supplementation was also recognized, and the addition of MOS tended to reduce fecal coliforms with an associated elevation in lactobacilli count compared to the control diet.
Summary
In summary, ‘biotic’ ingredients are becoming an increasingly popular inclusion within pet foods. It is evident there are great marketing opportunities surrounding their inclusion, which is supported by scientific research reflecting the benefits of their use.
References
Barry, K.A., Wojcicki, B.J., Middlebos, I.S., Vester, B.M., Swanson, K.S., & Fahey, G.C. Jr. (2010). Dietary cellulose, fructooligosaccharides, and pectin modify fecal protein catabolites and microbial populations in adult cats. Journal of Animal Science, 88(9), 2978-2987. https://pubmed.ncbi.nlm.nih.gov/20495116/
Beynen, A.C., Baas, J.C., Hoekemeijer, P.E., Kappert, H.J., Bakker, M.H., Koopman, J.P., & Lemmens, A.G. (2002). Faecal bacterial profile, nitrogen excretion, and mineral absorption in healthy dogs fed supplemental oligofructose. Journal of Animal Physiology and Animal Nutrition, 86(9-10), 298-305.
Gibson, G.R., & Roberfroid, M.B. (1995). Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. Journal of Nutrition, 125(6), 1401-1412.
Grieshop, C., Flickinger, E., Bruce, K., Patil, A.R., Czarnecki-Maulden, G.L., & Fahey, G.C. Jr. (2004). Gastrointestinal and immunological responses of senior dogs to chicory and mannan-oligosaccharides. Archives of Animal Nutrition, 58(6), 483-494.
Howard, M.D., Gordon, D.T., Garleb, K.A., & Kerley, M.S. (1995). Dietary fructooligosaccharide, xylooligosaccharide, and gum arabic have variable effects on cecal and colonic microbiota and epithelial cell proliferation in mice and rats. Journal of Nutrition, 125(10), 2604-2609.
Jenkins, D.J.A., Kendall, C.W.C., & Vuksan, V. (1999). Inulin, oligofructose, and intestinal function. Journal of Nutrition, 129(7), 1431S-1433S.
Kerry. (2020). Digestive health demands for pets are growing. Are your products ready? https://www.kerry.com/products/animal-applications/pet-food-nutrition/pet-digestive-health-ingredients
Kore, K.B., Pattanaik, A.K., Das, A., & Sharma, K. (2012). Evaluation of mannanoligosaccharide as prebiotic functional food for dogs: Effect on nutrient digestibility, hind gut health, and plasma metabolic profile. Indian Journal of Animal Sciences, 82(1), 81-86.
Pinna, C., Vecchiato, G., Bolduan, C., Grandi, M., Stefanelli, C., Windisch, W., Zaghini, G., & Biagi, G. (2018). Influence of dietary protein and fructooligosaccharides on fecal fermentative end-products, fecal bacterial populations, and apparent total tract digestibility in dogs. BMC Veterinary Research, 14, 106-115.
Propst, E.L., Flickinger, E.A., Bauer, L.L., Merchen, N.R., & Fahey, G.C. Jr. (2003). A dose-response experiment evaluating the effects of oligofructose and inulin on nutrient digestibility, stool quality, and fecal protein catabolites in healthy adult dogs. Journal of Animal Science, 81(12), 3057-3066.
Swanson, K.S., Grieshop, C.M., Flickinger, E.A., Bauer, L.L., Healy, H.P., Dawson, K.A., Merchen, N.R., & Fahey, G.C. Jr. (2002a). Supplemental fructooligosaccharides and mannanoligosaccharides influence immune function, ileal and total tract digestibilities, microbial populations, and concentrations of protein catabolites in the large bowel of dogs. Journal of Nutrition, 132(5), 980-989.
Swanson, K.S., Grieshop, C.M., Flickinger, E.A., Bauer, L.L., Chow, J., Wolf, B.W., Garleb, K.A., & Fahey, G.C. Jr. (2002b). Fructooligosaccharides and Lactobacillus acidophilus modify gut microbial populations, total tract nutrient digestibilities, and fecal protein catabolite concentrations in healthy adult dogs. Journal of Nutrition, 132(12), 3721-3731.
Swanson, K.S., Grieshop, C.M., Flickinger, E.A., Healy, H.P., Dawson, K.A., Merchen, N.R., & Fahey, G.C. Jr. (2002c). Effects of supplemental fructooligosaccharides plus mannanoligosaccharides on immune function and ileal and fecal microbial populations in adult dogs. Archives of Animal Nutrition, 56(5), 309-318.
Thompson, J.S., Quigley, E.M., Palmer, J.M., West, W.W., & Adrian, T.E. (1996). Luminal short-chain fatty acids and postresection intestinal adaptation. Journal of Parenteral and Enteral Nutrition, 20(5), 338-343.

Sophia Parkinson
GA Pet Food Partners Pet Food Claims Expert
Sophia Parkinson is GA Pet Food Partners' Pet Food Claims Expert, involved in checking partner claims, ensuring their labels and marketing materials meet regulations, and researching new and exciting raw materials. Sophia has an undergraduate degree in Nutritional Sciences, where she developed a strong interest in claims and labeling regulation. She briefly worked in the human food industry before joining GA in 2020. In her spare time, she enjoys cooking and going for long walks with her miniature schnauzer, Dexter.
You may also like...



Article written by Sophia Parkinson